RESUMEN
El objetivo de este estudio fue investigar los efectos de algunas hormonas vegetales en la germinación de semillas de Carica papaya L. del grupo “Solo” extraídas de frutos en etapas de madurez 3 y 5 (50% y 75% de la piel del fruto mostrando un color amarillo color). Luego, las semillas fueron sometidas a tratamientos que involucraron diferentes combinaciones de ácido 2-cloroetilfosfónico – (CEPA 0; 5 x 10-7 y 5 x 10 -4 M), GA 3 (0; 10 y 50 mg / L) y KNO 3 (0 o 1 M), y se evaluó la germinación a los 14 y 30 días. Las semillas de frutos en la etapa de madurez 3 mostraron una mayor germinación cuando se trataron con todos los reguladores de crecimiento probados. KNO 3promovió una disminución en la germinación y en el índice de velocidad de germinación, especialmente en las semillas de la etapa 5. Las hormonas vegetales no promovieron ningún aumento en el porcentaje de plántulas normales de semillas de papaya. El ácido dos-cloetilfosfónico (5 x 10 -4 M) y el ácido giberélico aplicado solo en las semillas de satge 3 promovieron un aumento en el número de plántulas normales 14 días después de la siembra.
Términos del índice: Carica papaya , CEPA, AG, fitohormonas, KNO 3 .
CURRÍCULUM
Las semillas de papaya tienen una germinación lenta y desigual, lo que se ha atribuido a la latencia poscosecha. El objetivo de este estudio fue evaluar los efectos de los reguladores de crecimiento sobre la germinación de semillas de papaya en el grupo “Solo”. Las semillas se extrajeron de frutos en las etapas de maduración 3 y 5, que corresponden al 50% y más del 75% de la superficie externa amarilla, respectivamente. Luego, fueron sometidos a tratamientos consistentes en diferentes combinaciones de ácido 2-cloroetilfosfónico – CEPA (0; 5 x 10-7 y 5 x 10 -4 M), GA 3 (0; 10 y 50 mg / L) y KNO 3(0 o 1 M), evaluando la germinación a los 14 y 30 días. Hubo un aumento en la germinación de semillas en la etapa 3 de maduración cuando se trataron con los reguladores de crecimiento probados. El KNO 3 tuvo un efecto negativo en la germinación y en el índice de velocidad de germinación, principalmente en las semillas de la etapa 5. No hubo efecto de los reguladores del crecimiento en el porcentaje final de plántulas normales. CEPA (5 x 10 -4 M) y las giberelinas utilizadas solas promovieron un aumento en el número de plántulas normales de semillas en la etapa 3, a los 14 días después de la siembra.
Términos del índice: Carica papaya , CEPA, giberelina, reguladores del crecimiento, KNO 3 .
Introducción
La germinación de las semillas de papaya ( Carica papaya ) es lenta y variable (Tokuhisa et al., 2007a), lo que puede atribuirse a la acción de compuestos fenólicos (Tokuhisa et al., 2007b). Algunos estudios demostraron un efecto positivo de las hormonas vegetales sobre la germinación de estas semillas, aumentando la velocidad de germinación y la uniformidad de las semillas de papaya del grupo “Formosa” (Tokuhisa et al., 2007a; Lopes et al., 2009). Sin embargo, los estudios sobre semillas del grupo “Solo” son escasos.
Entre los reguladores del crecimiento que influyen en la germinación de las semillas de papaya se encuentran las giberelinas (Tokuhisa et al., 2007a; Lopes et al., 2009), etileno (Linkies et al., 2009) y KNO 3.(Tokuhisa et al., 2007a; Dias et al., 2012). Las giberelinas actúan en la movilización de las reservas de semillas durante el proceso de germinación (Müller et al., 2006). Por lo tanto, se consideran importantes promotores de la germinación y contribuyen a aumentar la velocidad y uniformidad de la germinación de las semillas, mejorando así el rendimiento de las semillas de papaya (Tokuhisa et al., 2007a; Lopes et al., 2009). El etileno también se considera un promotor de la germinación de semillas. Esta hormona inhibe la acción del ácido abscísico y provoca el debilitamiento y ruptura del tegumento (Linkies et al., 2009). Para liberar etileno artificialmente en las células, se usa ácido 2-cloroetilfosfónico – CEPA (Burin et al., 1987).
El KNO3 es otro importante promotor de la germinación de semillas de papaya (Tokuhisa et al., 2007a; Dias et al., 2012). Sin embargo, hasta el momento no se ha entendido completamente su efecto específico porque puede actuar sobre la biosíntesis y / o una mayor sensibilidad a las giberelinas (Gb) (Alboresi et al., 2005).
Otro hecho que puede afectar el porcentaje y la velocidad de germinación de las semillas de papaya es la etapa de madurez del fruto (Lopes et al., 2009). Las semillas extraídas de frutos en diferentes etapas de madurez presentan diferentes niveles de sensibilidad a los reguladores del crecimiento (Firn, 1986). Sin embargo, existe poca información sobre la relación de la germinación de las semillas de papaya con la acción de los reguladores del crecimiento y la etapa de madurez del fruto.
Así, el presente trabajo tuvo como objetivo evaluar los efectos de CEPA, GA 3 y KNO 3 sobre la germinación de semillas de papaya “Solo” recolectadas de frutos en dos etapas de madurez, a saber, las etapas 3 y 5.
Material y métodos
El estudio se realizó en el Laboratorio de Análisis de Semillas del Departamento de Ciencias Vegetales de la Universidad Federal de Viçosa, Viçosa, MG, Brasil, utilizando semillas de papaya “Solo”. Los frutos se recolectaron en la etapa 3 de madurez (semi-madura, es decir, 50% de piel amarilla) y etapa 5 (madura, más del 75% de piel amarilla superficial), según Lopes et al. (2009).
Una vez extraídas de los frutos, las semillas se frotaron con hojas de papel y se lavaron con agua corriente para eliminar la sarcotesta. Luego, se secaron naturalmente sobre tres hojas de papel toalla y se mantuvieron en condiciones de laboratorio hasta alcanzar un contenido de humedad de aproximadamente el 10%, lo que ocurrió en aproximadamente 10 días. El contenido de agua de las semillas se determinó por el método del horno a 105 ± 3 o C durante 24 horas, con dos repeticiones (Brasil, 2009) de 50 semillas cada una. Los resultados se expresaron en porcentaje.
Después del secado, las semillas se trataron con hormonas vegetales. Para lograr esto, las semillas se sumergieron primero en agua destilada o en una solución de 1 M de KNO 3 durante 60 min y luego se lavaron con agua corriente. Posteriormente, se realizó la prueba de germinación de las semillas, la cual se realizó en papel toalla humedecido con las siguientes soluciones:
– Ácido giberélico (GA 3 ): en la concentración de 0 (agua destilada), 10 mg / L (2.8 x 10-5 M) y 50 mg / L (1.4 x 10 -4 M);
– CEPA: en la concentración de 0 (agua destilada), 5 x 10-7 M y 5 x 10 -4 M.
El tratamiento control consistió en semillas sumergidas únicamente en agua destilada durante 60 minutos y puestas a germinar en papel toalla humedecido con agua destilada.
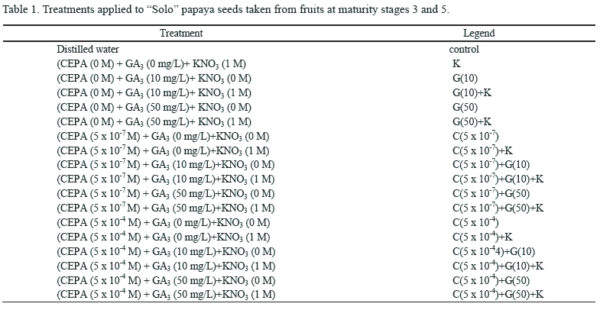
Los tratamientos experimentales utilizados se muestran en la Tabla 1 .
Para la prueba de germinación se colocaron cuatro réplicas de 50 semillas por cada etapa de madurez y para cada tratamiento sobre dos hojas de papel toalla humedecidas con un volumen de solución (agua para el control o CEPA + GA 3 + KNO 3), equivalente a 2,5 veces la masa del papel seco. Luego, las semillas se cubrieron con otra hoja de papel toalla y se hicieron los rollos. Los rollos se mantuvieron en un germinador tipo Mangelsdorff a una temperatura alterna de 20-30 ºC (16/8 horas, noche / día, respectivamente), según Brasil (2009). El índice de germinación se registró cada dos días durante 30 días. Las semillas que presentaban protuberancias radicales se consideraron germinadas. Se consideró plántulas normales aquellas que tengan la parte aérea de al menos 2 cm de largo. Se registraron los porcentajes de semillas que mostraron emergencia de radícula a los 14 días y 30 días de siembra, así como los porcentajes de plántulas normales (Brasil, 2009). Los datos obtenidos cada dos días se utilizaron para estimar la tasa de velocidad de protrusión de la radícula (RPSR) con base en los datos de emergencia de la raíz primaria,
El experimento se llevó a cabo en un diseño de bloques completos al azar, con cuatro repeticiones. Los datos fueron sometidos al análisis de varianza en un diseño factorial de 2 x 18 correspondientes a dos etapas de madurez y 18 combinaciones de reguladores de crecimiento. Para efectos de análisis, los datos porcentuales se transformaron en arc-sen 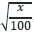 , debido a que no habían presentado distribución normal por la prueba de Kolmogorov-Smirnov. Sin embargo, en las tablas se expresaron los primeros valores. Los promedios del tratamiento, es decir, las diferentes concentraciones de reguladores del crecimiento, se compararon mediante la prueba de Scott-Knott. Las medias relativas a las etapas de madurez se compararon mediante la prueba F, con una probabilidad del 5%. Los análisis estadísticos se realizaron utilizando el paquete estadístico SAS.
, debido a que no habían presentado distribución normal por la prueba de Kolmogorov-Smirnov. Sin embargo, en las tablas se expresaron los primeros valores. Los promedios del tratamiento, es decir, las diferentes concentraciones de reguladores del crecimiento, se compararon mediante la prueba de Scott-Knott. Las medias relativas a las etapas de madurez se compararon mediante la prueba F, con una probabilidad del 5%. Los análisis estadísticos se realizaron utilizando el paquete estadístico SAS.
Resultados y discusión
En el día 14 después de la siembra, para la etapa de madurez 3, el porcentaje de semillas de control (no tratados con hormonas vegetales) que presentó emergencia de la radícula era del 66%, mientras que el porcentaje de plántulas normales correspondió a 18%. Para la etapa 5, el 73% de las semillas mostró emergencia de radícula y el 32% de plántulas normales ( Cuadro 2 ).

Las semillas de papaya de la etapa de madurez 3, 14 días después de la siembra, presentaron un aumento de 15 puntos porcentuales en el porcentaje de plántulas normales, en comparación con las semillas control, cuando fueron tratadas con CEPA 5 x 10 -4 M más KNO 3 . Las semillas en etapa de madurez 5 no se vieron afectadas por CEPA cuando se trataron simultáneamente con KNO 3 . Pero sin KNO 3 hubo una reducción de la germinación de hasta 25 puntos porcentuales en comparación con las semillas de control ( Cuadro 2 ).
Las semillas en etapa de madurez 3 tratadas con CEPA (5 x 10 -4 M) y GA 3 mostraron un aumento significativo en el porcentaje de plántulas normales en comparación con las semillas control, 14 días después de la siembra ( Cuadro 2 ). El papel de estos reguladores (etileno y giberelina) en la germinación de las semillas está asociado al debilitamiento de los tejidos que recubren al embrión, como el endospermo, la capa de aleurona y la frente (Linkies et al., 2009). Entonces, se puede inferir que el efecto de estos reguladores permitió una mayor germinación de las semillas de papaya.
Los tratamientos con CEPA en una concentración 5 x 10 -7 M causaron un efecto negativo en la producción de plántulas normales 14 días después de la siembra para las semillas de ambas etapas de madurez. Se encontró una disminución de 18% a 5% para las semillas de la etapa 3 y de 32% a 7% para las semillas de la etapa 5, en comparación con las semillas de control ( Cuadro 2 ). En la literatura se encuentran diferentes efectos del etileno en la germinación de semillas, y esta hormona vegetal, junto con GA o citoquinina, provocó una disminución en la tasa de germinación de la maracuyá dulce (Zucareli et al., 2003); el etileno no afectó la germinación de semillas de arroz rojo, asociado o no con inhibidores (2-aminoetoxivinilglicina y tiosulfato de plata) (Gianinetti et al., 2007); y resultó ser clave para superar el letargo deStylosanthes humilis (Ribeiro y Barros, 2006) .
No hubo diferencia en el porcentaje de semillas con protuberancia radícula y en el porcentaje de plántulas normales, medido 30 días después de la siembra, entre las semillas de ambas etapas de madurez y las que no recibieron tratamiento. Correspondieron al 71% y 81% para las semillas de los estadios 3 y 5, respectivamente. El porcentaje de plántulas normales fue de 71% y 75% para las semillas de las etapas 3 y 5, respectivamente ( Cuadro 3 ).
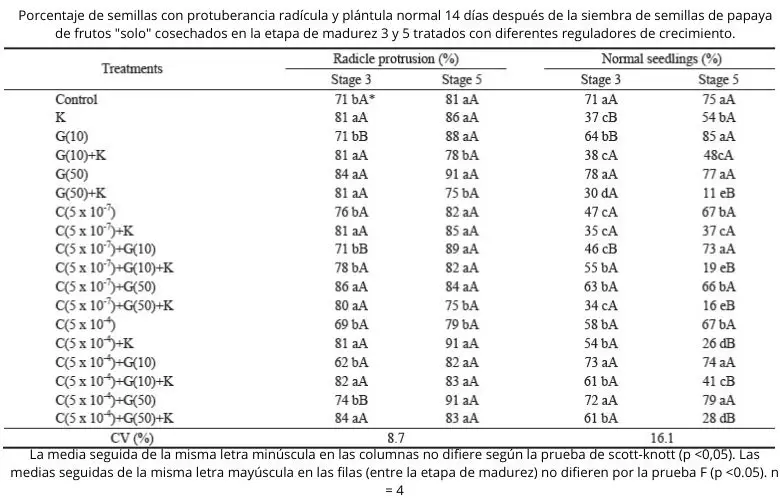
Se sabe que los AG aumentan el índice de germinación de las semillas de papaya (Tokuhisa et al., 2007a). GA 3 solo, en el 14 ° día después de la siembra, casi se duplicó el número de plántulas normales para la etapa 3 semillas, es decir, se incrementó de 18% a 38%, en comparación con semillas no tratadas. Por otro lado, GA 3 provocó una reducción de siete puntos porcentuales en el porcentaje de plántulas normales de semillas de la etapa 5 en comparación con las semillas sin tratar.
Las semillas de la etapa 3 presentaron un mayor porcentaje de plántulas normales en comparación con las semillas de la etapa 5 cuando se trataron con GA 3 (10 mg / L) ( Cuadro 2 ). Según Lopes et al. (2009), las semillas de papaya del grupo “Formosa” extraídas de frutos en etapa de madurez 3 presentaron porcentajes más altos de plántulas normales cuando fueron tratadas con GA 3, en comparación con semillas en otras etapas de madurez. Así, la sensibilidad a los reguladores de crecimiento es diferente en cada etapa de madurez de las semillas, lo que puede presentar una mayor o menor sensibilidad a los reguladores utilizados (Firn, 1986; Alberts et al., 2010).
Según Santos et al. (1999), semillas recién recolectadas de papaya del grupo “Solo” presentaron máxima germinación en la etapa de madurez 5. El porcentaje de plántulas normales de semillas control, 30 días después de la siembra, fue casi 73% ( Cuadro 3 ). Martins et al. (2006) y Carlesso et al. (2009).
El nitrato de potasio provocó un aumento significativo de la germinación de las semillas de la etapa 3, correspondiente a 18 puntos porcentuales, 14 días después de la siembra y 10 puntos porcentuales 30 días después de la siembra, en comparación con la germinación de semillas sin tratar. Pero, a pesar de presentar la raíz primaria, la mayoría de las semillas no dieron como resultado plántulas normales en presencia de KNO 3 . Se encontró una reducción del 48% y 28% en plántulas normales para las semillas de la etapa 3 y 5, respectivamente, en comparación con las semillas sin tratar ( Tablas 2 y 3 ).
Meireles y col. (2009) también observaron un efecto negativo del KNO 3 en la germinación de semillas de papaya. Según este autor, una mayor concentración de potasio en el agua durante el riego resultó en una disminución del 30% en la germinación de la semilla. Además, Tokuhisa et al. (2007a) encontraron reducción en el porcentaje de germinación de semillas de papaya “Formosa”, que no presentaron latencia, sumergidas en una solución de nitrato de potasio por 120 minutos en una concentración de 1 M. Bhattacharya y Khuspe (2001) también observaron que semillas de papaya que habían sido previamente sumergidas en KNO 3 0.1 M durante 24 horas presentó una reducción en el porcentaje de plántulas normales. El número reducido de plántulas normales encontradas a partir de semillas tratadas con KNO 3pudo haber sido causado por estrés hídrico y / o estrés salino, al que las semillas habían estado expuestas, una vez que la concentración de 1 M utilizada, durante 60 minutos, pudo haber causado daños a las semillas. La disminución del potencial hídrico puede haber estado asociada con la reducción de la disponibilidad y difusión de oxígeno en la solución de imbibición (Lopes y Souza, 2008).
Los efectos negativos del KNO 3 en el número de plántulas normales, 30 días después de la siembra, fueron causados por GA 3 (50 mg / L), con una reducción del 42% (etapa 3) y 64% (etapa 5) en el porcentaje de plántulas normales, en comparación con las semillas de control ( Cuadro 3 ).
El tratamiento con KNO 3 resultó en un aumento del índice de velocidad de emergencia (ESI) de las semillas de ambas etapas de madurez. Sin embargo, el índice de velocidad de germinación (GSI) fue menor cuando se trató con esta sal. Las semillas de la etapa 5 fueron más sensibles a la solución de KNO 3 , presentando una mayor reducción en el porcentaje de plántulas normales. En general, la aplicación de reguladores de crecimiento en las semillas de la etapa 3, a excepción de CEPA 5 x 10 -7 M sin KNO3 (Nitrato de potasio) , resultó en un aumento del ESI de las semillas ( Cuadro 4 ).
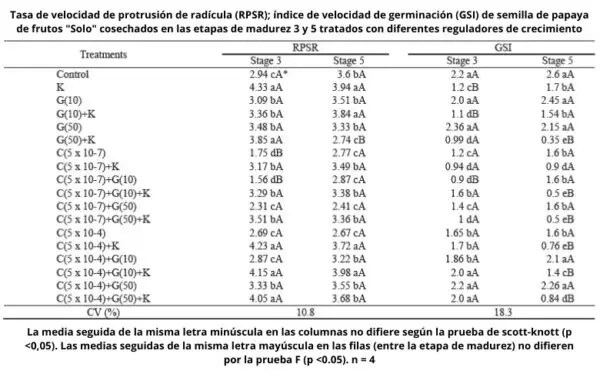
Los reguladores del crecimiento como GA 3 y nitrato de potasio fueron efectivos para aumentar la velocidad de germinación y también para estimular la germinación de semillas de papaya (Tokuhisa et al., 2007a; Lopes y Souza, 2008; Lopes et al., 2009), incluso con una tasa reducida de plántulas normales ( Cuadro 4 ).
Conclusiones
Hay un aumento en el índice de germinación de las semillas en etapa de madurez 3 cuando se tratan con las hormonas vegetales probadas. El KNO 3 tiene un efecto negativo sobre la germinación y sobre el índice de velocidad de germinación, especialmente en las semillas de la etapa 5. No hay efecto de las hormonas vegetales en el porcentaje de plántulas normales. CEPA (5 x 10 -4 M) y las giberelinas empleadas solas provocaron un aumento en el número de plántulas normales de las semillas de la etapa 3, 14 días después de la siembra.
Referencias
ALBERTS, B.; JOHNSON, A.; LEWIS, J.; RAFF, M.; ROBERTS, K.; WALTER, P. Biología molecular de la célula . 5.ed. Porto Alegre: Artmed Editora, 2010. 1396 p. [ Enlaces ]
ALBORESI, A.; GESTIN, C.; LEYDECKER, MT.; BEDU, M.; MEYER, C.; TRUONG, HN. Nitrato, una señal que alivia la latencia de las semillas en Arabidopsis. Plant Cell and Environment , v.28, n.4, p.500 – 512, 2005. http://www.ncbi.nlm.nih.gov/pubmed/16229082
BHATTACHARYA, J.; KHUSPE, SS Germinación in vitro e in vivo de semillas de papaya ( Carica papaya L.). Scientia Horticulturae , v.91, n.1, p.39-49, 2001. http://www.sciencedirect.com/science/article/pii/S0304423801002370
BRASIL. Ministerio de Agricultura, Ganadería y Abastecimiento. Reglas para el análisis de semillas . Ministerio de Agricultura, Ganadería y Abastecimiento. Secretaría de Defensa Agraria. Brasilia: MAPA / ACS.2009. 395p. http://www.bs.cca.ufsc.br/publicacoes/regras%20analise%20sementes.pdf [ Enlaces ]
BURIN, YO; BARROS, RS; RENA, AB Regulación química de la latencia endógena en semillas de Stylosanthes humilis HBK Turrialba , v.37, p.281-285, 1987. [ Enlaces ]
CARLESSO, VO; BERBERT, PA; SILVA, RF; THIÉBAUT, JTL; OLIVEIRA, MTR Germinación y vigor de papaya ( Carica papaya L.) cv. Dorado secado a altas temperaturas. Revista Brasileira de Sementes , v.31, n.2, 228-235, 2009. http://www.scielo.br/pdf/rbs/v31n2/v31n2a27.pdf [ Links ]
DIAS, MA; SOUZA NETO, JD; CONCEIÇÃO, PM; TOREZANI, SC Respuesta fisiológica de semillas de papaya sometidas a acondicionamiento osmótico. Revista Caatinga , vol. 25, n. 4, pág. 82-87, 2012. http://200.137.6.4/revistas/index.php/sistema/article/view/2389 [ Enlaces ]
FIRN, RD Sensibilidad a las sustancias de crecimiento: la necesidad de ideas más claras, términos de necesidad y experimentos con propósito. Physiologia Plantarum , v.67, n.2, p.267-272, 1986. [ Enlaces ]
GIANINETTI, A.; LAARHOVEN, LJJ; PERSIJN, ST; HARREN, FJM; PETRUZZELLI, L. La producción de etileno está asociada con la germinación pero no con la latencia de las semillas en el arroz rojo. Annals of Botany , v.99, n. 4, p. 735-745, 2007. http://aob.oxfordjournals.org/content/99/4/735.full.pdf+html [ Enlaces ]
LINKIES, A.; MÜLLER, K.; MORRIS, K.; TURECˇKOVA´, V.; CADMAN, CSC; CORBINEA, UF; STRNAD, M.; LYNN, JR; FINCH-SALVAJE, NOSOTROS; LEUBNER-METZGER, G. El etileno interactúa con el ácido abscísico para regular la ruptura del endospermo durante la germinación: un enfoque comparativo utilizando Lepidium sativum y Arabidopsis thaliana . The Plant Cell , v.21, n.12, p.3803-3822, 2009. http://www.dbbe.fcen.uba.ar/new/contenido/objetos/endospermagerminacion.pdf [ Links ]
LOPES, HM; SOUZA, CM Efectos de la giberelina y el secado sobre el acondicionamiento osmótico sobre la viabilidad y vigor de las semillas de papaya ( Carica papaya L.). Revista Brasileira de Sementes , v.30, n.1, p.181-189, 2008. http://www.scielo.br/pdf/rbs/v30n1/a23v30n1.pdf [ Links ]
LOPES, AWP; SELEGUINI, A.; BOLIANI, AC; CÔRREA, LS Etapa de maduración del fruto y uso del ácido giberélico en la germinación de semillas de papaya. Pesquisa Agropecuária Tropical , v.39, n.4, p.278-284, 2009. http://www.revistas.ufg.br/index.php/pat/article/view/5763 [ Links ]
MAGUIRE, JD Velocidad de germinación: ayuda a la emergencia y vigor de las plántulas. Crop Science , v.2, n.2, p.176-177, 1962. [ Enlaces ]
MARTINS, GN; SILVA, RF; PEREIRA, MG; ARAÚJO, EF; POSSE, SCP La influencia de la fruta poscosecha se basa en la calidad fisiológica de las semillas de papaya. Revista Brasileira de Semente , v.28, n.2, p.142-146, 2006. http://www.scielo.br/pdf/rbs/v28n2/a19v28n2.pdf [ Links ]
MEIRELES, RC; SILVA, RF; ARAÚJO, EF; REIS, LS; LYRA, GB; MARINHO, A. Influencia del nitrógeno y las profundidades de riego en la calidad fisiológica de las semillas de papaya. Revista Brasileira de Sementes , v.31, n.1, p.216-221, 2009. http://www.scielo.br/pdf/rbs/v31n1/a24v31n1.pdf [ Links ]
MÜLLER, K.; TINTELNOT, S.; LEUBNER-METZGER, G. Germinación de semillas de Brassicaceae con endospermo limitado: el ácido abscísico inhibe el debilitamiento del endospermo inducido por el embrión de Lepidium sativum (berro) y la ruptura del endospermo del berro y Arabidopsis thaliana . Plant Cell Physiology , v.47, n. 7 p. 864-877, 2006. http://pcp.oxfordjournals.org/content/47/7/864.full.pdf+html [ Enlaces ]
RIBEIRO, DM; BARROS, RS Sensibilidad al etileno como componente principal en la germinación de semillas de Stylosanthes humilis. Seed Science Research , v.16, n.1, p.37-45, 2006. http://journals.cambridge.org/action/displayAbstract?fromPage=online&aid=705764 [ Enlaces ]
SANTOS, RCA; SAMPAIO, LSV; COSTA, JA Condición ambiental, contenido de agua y empaque sobre la viabilidad y vigor de las semillas de papaya. Revista Brasileira de Sementes , v.21, n.2, p.194-202, 1999. http://www.abrates.org.br/revista/artigos/1999/v21n2/artigo30.pdf [ Links ]
TOKUHISA, D.; DIAS, DCFS; ALVARENGA, EM; DIAS, LAS; MARIN, SLD Tratamientos para superar la latencia en semillas de papaya. Revista Brasileira de Sementes , v.29, n.1, p.131-139, 2007a. http://www.scielo.br/pdf/rbs/v29n1/18.pdf [ Enlaces ]
TOKUHISA, D.; DIAS, DCFS; ALVARENGA, EM; HILST, PC; DEMUNER, AJ Compuestos fenólicos que inhiben la germinación en semillas de papaya ( Carica papaya l.). Revista Brasileira de Sementes , v.29, n.3, p.180-188, 2007b. http://www.scielo.br/pdf/rbs/v29n3/a22v29n3.pdf [ Enlaces ]
ZUCARELI, C.; CASTRO, MM; OLIVEIRA, HR; BRANCALIÃO, SR; RODRIGUES, JD; ONO, EO; BOARO, CSF Fitorreguladores y germinación de semillas de maracuyá dulce en condiciones de laboratorio. Scientia Agrária , v.4, n.1-2, p.9-14, 2003. http://dialnet.unirioja.es/servlet/articulo?codigo=2910474 [ Links ]
¿Alguna queja o sugerencia?: Reportar